
Investigations dendrochronologiques au cœur de la toundra sibérienne
Sibérie - Toundra - Goulag
Dans un contexte fortement marqué par la hausse des températures à la surface du globe, l’Arctique représente une région où elles ont augmenté deux fois plus vite que dans le reste du monde durant les quarante dernières années. Cette augmentation est particulièrement rapide depuis 2010, atteignant 0.75°C entre 2010 et 2019 (Cohen et al., 2020 ; Overland, Wang, Walsh, & Stroeve, 2014 ; Post et al., 2019). Le réchauffement de l’Arctique et ses conséquences sur les écosystèmes font l’objet de multiples études mesurant les impacts sur la perte d’habitats pour les espèces endémiques, sur les changements de couverture végétale et sur l’évolution du pergélisol (Bjorkman et al., 2020 ; Mekonnen, Riley, Randerson, Grant, & Rogers, 2019 ; Post et al., 2019 ; Yletyinen, 2019).
En 2019, une expédition scientifique interdisciplinaire russo-suisse effectue une campagne de terrain dans le Grand Nord russe au sud de la péninsule du Yamal. La zone comprend un ancien camp du Goulag, situé le long de la "Voie morte", le camp 91 du chantier 501, dénommé Chtchoutchi.
La situation géographique du camp de Chtchoutchi révêt un intérêt particulier pour l’étude de la végétation présente et notamment les peuplements forestiers. En effet, l’étude des écosystèmes arctiques situés dans un écotone forêt-toundra est intéressante car ils sont particulièrement sensibles aux changements induits par le réchauffement climatique. La végétation fonctionne en système interconnecté complexe. De plus, à hautes latitudes, les conséquences biologiques des changements climatiques sont plus facilement mesurables en raison de la grande variation des températures observées depuis trois décennies. L’émergence de nouvelles techniques de télédétection a permis d’observer un "verdissement" de l’Arctique, indicateur de changement majeur (Myers-Smith et al., 2020 ; Nitze & Grosse, 2016).
La présente étude est exploratoire, elle tente d’évaluer les relations entre variables climatiques, les activités anthropiques et la croissance des arbres. En prélevant des carottes d’arbres directement sur place, plusieurs chronologies ont pu être construites, permettant d’évaluer l’impact d’une sélection de variables sur la croissance des arbres échantillonnés.
Contexte géographique
La zone d’étude se situe à la frontière entre deux aires biotiques terrestres majeures ; la toundra et la taïga. Le biome de la toundra consiste en une région de haute latitude ou de haute altitude au-dessus de la limite supérieure de la forêt ; avec une température annuelle moyenne inférieure à 0°C (Thomas et al., 2020), des quantités de précipitations et une activité éolienne trop importantes pour qu’une végétation forestière naturelle puisse se développer. Bien que la végétation de la toundra varie fortement à l’échelle locale et régionale (Walker et al., 2006), elle est principalement composée d’arbustes nains, d’herbacées (Carex), de mousses et de lichens. Plus humide et avec des températures légèrement plus élevées, la taïga représente quant à elle le biome sibérien spatialement le plus important. Le climat de la taïga est généralement froid avec une température moyenne négative plus de six mois par année. Elle se compose surtout de conifères, de mousses, d’arbustes et de lichens.
Notre site d’étude se trouve sur la surface marécageuse d’une plaine fluvio-lacustre dont l’altitude est comprise entre 25 m et 30 m au-dessus du niveau de la mer (Melnikov, Leibman, Moskalenko, & Vasiliev, 2004). Le sol est principalement composé de sable alluvial avec quelques couches argileuses.
Le climat local est semi-continental avec une température moyenne de -34°C en hiver et +10°C en été. La région est influencée par les masses d’air océaniques provenant de l’Arctique ainsi que continentales provenant de l’Asie, favorisant ainsi des conditions météorologiques très variables (Matyshak et al., 2017 ; NASA Earth Observatory, 2019). D’octobre à mars il fait normalement très froid, la couverture nuageuse est persistante, les chutes de neige sont intenses et entrecoupées de courtes périodes plus chaudes. Le printemps est plus lumineux, venteux, avec de faibles précipitations, alors que la période juin-juillet est souvent nuageuse, ponctuée de précipitations intenses mais sporadiques. Enfin, l’automne connaît une couverture nuageuse persistante avec des épisodes de pluie réguliers (Matyshak et al., 2017). Dans la région de Nadym, la température annuelle moyenne de l’air a augmenté d’environ +0,05°C par année entre 1960 et 2017 (Kukkonen et al., 2020), illustrant ainsi un réchauffement local rapide.
Pergélisol et végétation
La région du Yamal et celle de Nadym se trouvent sur du pergélisol. Ce dernier est défini comme un sol gelé en permanence, dont la température reste inférieure à 0°C pendant au moins deux années consécutives (Biskaborn et al., 2019).
Le pergélisol est composé d’une couche active, affectée par un dégel estival variable, puis d’une couche gelée en permanence. On retrouve du pergélisol sur 24% de l’hémisphère nord et environ 80% de la Sibérie (Tchebakova, Parfenova, & Soja, 2009). Notre site de recherche se situe à la limite entre une zone de pergélisol continu (le pergélisol est présent partout) et discontinu (l’occupation spatiale du pergélisol est interrompue par du sol non-gelé). L’épaisseur du pergélisol peut s’étendre de quelques mètres à plus de 700 mètres de profondeur selon le lieu (Singh et al., 2011). La couche active varie annuellement en fonction des conditions climatiques ; son épaisseur est également influencée par des facteurs tels que la couverture neigeuse, la couverture végétale, la topographie ou l’hydrologie (Tchebakova et al., 2009). En outre, des études ont montré que l’évolution de la température du pergélisol et l’évolution de sa fonte influencent la végétation présente au-dessus en fournissant davantage d’humidité au sol et favorisant par endroits l’expansion et la croissance des plantes (Frost & Epstein, 2014).
La végétation présente sur le site d’étude se compose surtout d’arbustes bas, d’herbacées (Carex), de mousses et lichens ainsi que de forêts clairsemées de bouleaux et mélèzes (Larix sibirica). Ces peuplements forestiers se situent principalement sur des terrains bien drainés, néanmoins certains peuplements présents dans des zones humides dépérissent par excès d’humidité et déstabilisation du sol. Cette végétation évolue rapidement ; en effet les travaux de télédétection que nous avons menés en analysant les images satellites nous ont montré une croissance significative de l’occupation du sol par des forêts éparses et des arbustes ainsi qu’une nette diminution des sols nus.
Méthode
Notre étude dendrochronologique s’est focalisée sur les mélèzes. Le Larix sibirica a une grande capacité d’adaptation. Il se développe sur des sites aux conditions environnementales différentes. Sensible à l'humidité du sol, le mélèze atteint rarement sa hauteur et sa largeur de tronc maximales si elle est trop élevée. Parmi les mélèzes, Larix sibirica est le plus sensible au pergélisol. Le manque d’air dans le sol et la basse température du sol entraînent une forte diminution de la productivité des peuplements (Abaimov, 2010). Dans notre zone d’étude, la période de croissance du mélèze dure environ 60 jours, soit entre la mi-juin et la mi-août (Devi et al., 2008 ; Osawa, Matsuura, & Kajimoto, 2010). La période de croissance dépend directement du dégel de la couche active, de la fonte du manteau neigeux et de la température de l’air. Cette dernière est d’ailleurs considérée comme le facteur limitant pour la croissance des arbres dans les régions de hautes latitudes (Antonova & Stasova, 1997). Cependant, depuis plusieurs années, la littérature remet en question l’importance de la température de l’air dans le processus de croissance des cernes des arbres (Briffa et al., 1998 ; D’Arrigo, Wilson, Liepert, & Cherubini, 2008 ; Kharuk, Ranson, Im, & Petrov, 2015 ; Wilmking, 2005).
À l’instar des autres conifères, le mélèze forme un cerne de croissance par année. Le cerne se compose de deux parties : un bois initial clair, de faible densité, à trachéides ("cellules") larges et parois minces, qui se forme en début de saison, et un bois final plus sombre, de densité supérieure avec des trachéides de petite taille, à parois épaisses, qui se forme en fin de saison (Corona, 2014). La taille des cernes varie en fonction des conditions environnementales. Dans notre cas, une courte période de croissance combinée à un environnement relativement froid provoque parfois le développement incomplet de cernes ou la formation de cerne très fin voire l’absence de cerne (Gurskaya, 2019). Parmi les échantillons étudiés, des cernes manquants ont régulièrement été relevés et la largeur des cernes varie entre 0.02 mm et 4.0 mm.
La zone d’étude a été divisée en 4 sites d’échantillonnage distincts autour du camp de base (AF, CT, GU et VF) (cf. StoryMap), permettant ainsi une meilleure couverture de la zone et la comparaison des résultats obtenus pour chacun des sites. Nous avons sélectionné les sites d’échantillonnage en fonction de leur accessibilité, de la présence des peuplements de mélèzes et de leur distance par rapport au site du goulag, et donc de l’activité anthropique potentielle passée. Chaque site répond à ces critères de manière différente. Un minimum de 30 arbres a été prélevé sur chaque site et chaque arbre échantillonné a été géoréférencé et photographié.
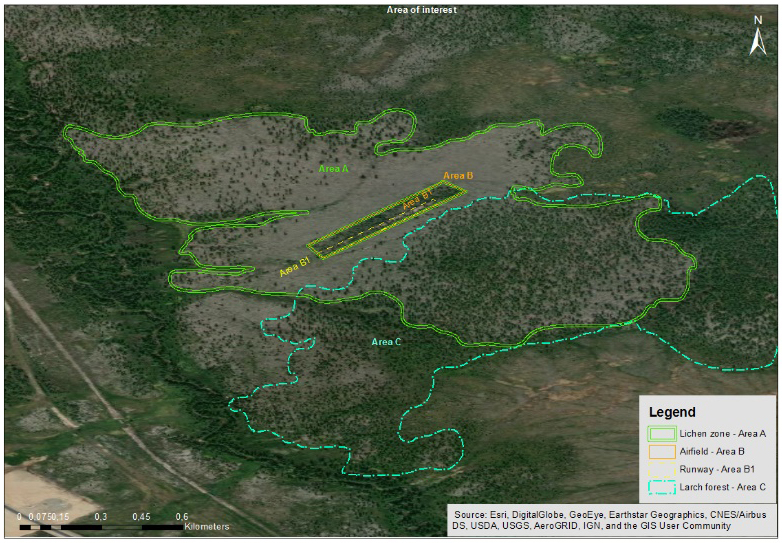
Figure 1 : Site AF, zone de la supposée piste pour aéronefs. Les frontières marquées par les espèces végétales sont clairement distinctes.
Le site AF est un site situé à environ 900 m au nord-est du camp (Figure 1). Au cours de l’expédition, l’historien Vadim Gritsenko a émis l’hypothèse que la zone pourrait avoir été utilisée comme terrain d’aviation pendant la période de l’Union soviétique. Un ancien aéronef accidenté se trouvant à proximité de la piste d'atterrissage supposée corrobore cette hypothèse. Le site AF est une zone plane et bien drainée, la couverture végétale est dominée par les lichens (C. rangiferina), elle comprend également des peuplements de mélèzes, de bouleaux et quelques pins (Pinus sibirica) isolés. La Figure 1 montre la piste de l’aérodrome couverte de mélèzes et de bouleaux. À l’extérieur du rectangle, la zone a été déboisée ; nous avons trouvé des souches avec des marques de haches et les troncs laissés à proximité. Malgré la dégradation des souches, nous avons pu échantillonner certaines d’entre-elles et estimer la date de coupe à l’année 1916. La stratégie d’échantillonnage s’est organisée le long de transects linéaires traversant le site du nord-ouest au sud-est.
L’étude de ce peuplement nous a permis d’obtenir des informations sur le lieu et notamment de dater l’âge des arbres présents sur la piste. Les résultats détaillés sont visibles dans la StoryMap qui illustre ce billet de blog.
Le site CT est situé à environ 12 km au nord-ouest du camp de base. Il s’agit d’une zone marécageuse difficile d’accès, comprenant des mélèzes, des pins, de la végétation naine et des mousses. Une grande majorité des arbres sont penchés en raison d’un excès d’humidité et donc de l’instabilité du sol. Ce site a été sélectionné à partir de l’observation des images satellite et des possibilités logistiques pour s’y rendre. Après discussion avec l’historien, il s’est avéré que ce site pouvait correspondre à une zone n’ayant probablement pas connu d’activité anthropique en lien avec l’activité du goulag, excluant ainsi l’influence de l’activité humaine sur la croissance des arbres. À l’exception d’une route construite à 400 m du site en 2018-2019, le site est resté vierge de toute activité anthropique. En outre, selon les informations recueillies par V. Gritsenko, les impacts anthropiques des camps du goulag s’étendaient généralement dans un rayon de 5 km autour du camp. Notre site dépassait donc largement ce rayon.
Le site GU correspond au camp de Chtchoutchi. Les arbres présents au moment de la construction ont tous été défrichés pour permettre la construction des infrastructures du camp. Nous nous attendons par conséquent à y trouver des arbres relativement jeunes, ayant poussé après l’abandon du camp en 1949. La végétation du site est composée de mélèzes ayant recolonisé le site et de deux allées de pins, plantées lorsque le camp était en activité, a priori à des fins d’embellissement du camp. L’âge de ces arbres est visible dans la StoryMap ; étant donné leur jeune âge, ce site a été exclu de l’analyse dendroclimatologique mais permet néanmoins de corroborer les informations sur l’abandon du camp vers 1951-1952.
Le site VF est situé à proximité du camp 93 (550 m) et de la voie ferrée. Les arbres ont été échantillonnés selon un transect perpendiculaire à la voie ferrée. Il s’agit d’une zone mixte de sol drainé et de terrain humide et marécageux. Le site a été relativement épargné par le défrichement ; les arbres forment des peuplements denses.
Résultats
Dendroclimatologie des sites AF, CT et VF
L’expédition in situ de 2019 nous a permis de prélever 109 carottes provenant des arbres sur les sites AF, CT et VF. Après le post-traitement de ces échantillons nous avons exclu 42 carottes. Parmi ces dernières, 21 provenaient d’arbres trop jeunes, ne permettant pas d’interprétation fiable des cernes de croissance ; 7 provenaient de pins, exclus pour uniformiser l’étude autour du mélèze ; et 14 n’ont malheureusement pas pu être interdatés. Grâce aux carottes exploitables, il a été possible de construire trois chronologies.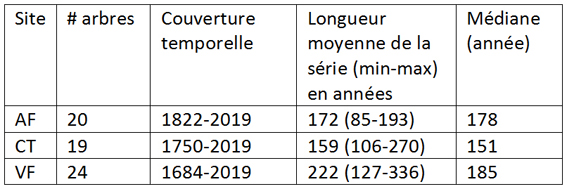
Tableau 1 : Récapitulatif des échantillons par site.
Après avoir standardisé les chronologies obtenues, nous avons pu comparer la croissance des arbres sur les trois sites. Bien que la tendance de croissance soit similaire sur les trois sites depuis les années 1830, le site AF montre une croissance accélérée depuis les années 2000, peut-être en lien avec un meilleur drainage naturel du terrain. Afin de vérifier la similarité de croissance dans ces chronologies, nous avons produit une matrice de corrélation. La corrélation croisée des carottes fait alors apparaître des groupes d’intercorrélation positive et quelques groupes mineurs d’intercorrélation négative. Ces résultats suggèrent que la croissance des arbres s’explique davantage par des facteurs régionaux plutôt que spécifiques aux sites. Ainsi, il nous a été possible de créer une chronologie unique pour cette région en intégrant nos trois chronologies au sein d’une seule chronologie régionale.
Relation entre climat et croissance des arbres
Nous avons identifié le facteur principal de croissance des arbres échantillonnés et nous avons déterminé qu’il s’explique à l’échelle régionale. Il est donc possible d’approfondir nos recherches sur les relations entre climat et croissance. Nous observons que pour la période 1901-2018, le coefficient de corrélation sur les trois sites est significativement positif par rapport aux températures des mois de juin et de juillet, alors qu’il est significativement négatif par rapport aux températures du mois d’avril. Les coefficients restent même significatifs avec un intervalle de confiance de 99%. L’observation de ces résultats nous permet de déduire que la croissance des arbres de notre zone d’étude dépend principalement de la température estivale, tout en étant moins sensible aux précipitations.
Les fonctions de corrélation utilisées montrent également une relation significative entre l’indice standardisé de précipitation et d’évapotranspiration (SPEI) et la croissance des arbres. En effet, les coefficients de corrélation sont positifs entre décembre de l’année précédente et le mois de mai de l’année en question ; et négatifs entre juin et novembre. Cette observation signifie que les arbres semblent répondre positivement à l’environnement humide durant les premiers mois de l’année, puis négativement durant l’été en raison d’une humidité trop élevée durant cette saison. La corrélation positive durant l’hiver suggère que le manteau neigeux pourrait avoir une fonction isolante et donc une influence sur la croissance des arbres en début d’année. Néanmoins, les résultats obtenus de l’indice de corrélation entre le SPEI et les cernes de croissance restent difficiles à interpréter.
Conclusion
La présente étude avait pour but initial d’évaluer les effets des activités anthropiques et de l’évolution du pergélisol sur la croissance des arbres, en analysant leurs cernes de croissance. La campagne menée sur le terrain nous a permis de disposer de nos propres échantillons d’arbres. Les arbres de cette région sont idéaux pour isoler l’influence de la température, facteur limitant principal de leur croissance. La campagne d’échantillonnage a permis de constituer trois chronologies de mélèzes centenaires. L’analyse de ces séries a mis en évidence les relations entre la croissance des cernes des arbres, la température et les précipitations, dans une région reconnue pour l’évolution rapide des changements climatiques. Il est intéressant de relever que la croissance des arbres est davantage influencée par les conditions climatiques locales que par les activités anthropiques passées.
Néanmoins, les sites où l’activité humaine a été plus marquée, à l’instar de l’aérodrome, du camp et de la voie de chemin de fer proche, ont vu leur sol et leur couverture végétale altérés. Si notre hypothèse de départ consistait à prédire que les arbres échantillonnés sur des sites différents présenteraient des tendances de croissance différentes, en fonction de leur proximité avec les zones impactées par les activités anthropiques, les analyses nous montrent le contraire, à quelques exceptions près. Nos données montrent que les différents sites étudiés sont positivement intercorrélés. Cependant, il semble que les populations locales sont plus impactées par les changements climatiques que par les activités humaines sporadiques, d’une ampleur relativement faible, ou par les facteurs environnementaux locaux tels que les sols ou l’humidité.
Liens
StoryMap ArcGIS : Expédition Changing Arctic 2019
Site de référence du projet : Yamal.ch
Références
Abaimov, A.P. (2010). "Geographical distribution and genetics of Siberian larch species". In Permafrost ecosystems (pp. 41-58). Springer.
Antonova, G.F., & Stasova, V.V. (1997). "Effects of environmental factors on wood formation in larch (Larix sibirica Ldb.) stems". Trees, 11(8), 462-468. doi.org/10.1007/PL00009687.
Biskaborn, B.K., Smith, S.L., Noetzli, J., Matthes, H., Vieira, G., Streletskiy, D.A., … Lantuit, H. (2019). "Permafrost is warming at a global scale". Nature Communications, 10(1), 264. doi.org/10.1038/s41467-018-08240-4.
Bjorkman, A.D., García Criado, M., Myers-Smith, I.H., Ravolainen, V., Jónsdóttir, I.S., Westergaard, K.B., … Normand, S. (2020). "Status and trends in Arctic vegetation: Evidence from experimental warming and long-term monitoring". Ambio, 49(3), 678-692. doi.org/10.1007/s13280-019-01161-6.
Briffa, K.R., Schweingruber, F.H., Jones, P.D., Osborn, T.J., Shiyatov, S.G., & Vaganov, E.A. (1998). "Reduced sensitivity of recent tree-growth to temperature at high northern latitudes". Nature, 391(6668), 678-682. doi.org/10.1038/35596.
Cohen, J., Zhang, X., Francis, J., Jung, T., Kwok, R., Overland, J., … Yoon, J. (2020). "Divergent consensuses on Arctic amplification influence on midlatitude severe winter weather". Nature Climate Change, 10(1), 20-29. doi.org/10.1038/s41558-019-0662-y.
Corona, C. (2014). La dendrogéomorphologie.
D’Arrigo, R., Wilson, R., Liepert, B., & Cherubini, P. (2008). "On the 'Divergence Problem' in Northern Forests: A review of the tree-ring evidence and possible causes". Global and Planetary Change, 60(3-4), 289-305. doi.org/10.1016/j.gloplacha.2007.03.004.
Devi, N., Hagedorn, F., Moiseev, P., Bugmann, H., Shiyatov, S., Mazepa, V., & Rigling, A. (2008). "Expanding forests and changing growth forms of Siberian larch at the Polar Urals treeline during the 20th century". Global Change Biology, 14(7), 1581-1591. doi.org/10.1111/j.1365-2486.2008.01583.x.
Frost, G.V., & Epstein, H.E. (2014). "Tall shrub and tree expansion in Siberian tundra ecotones since the 1960s". Global Change Biology, 20(4), 1264-1277. doi.org/10.1111/gcb.12406.
Gurskaya, M.A. (2019). "Effect of Summer Monthly Temperatures on Light Tree Ring Formation in Three Larch Species (Larix) in the Northern Forest-Tundra of Siberia". Russian Journal of Ecology, 50(4), 343-351. doi.org/10.1134/S1067413619040088.
Kharuk, V.I., Ranson, K.J., Im, S.T., & Petrov, I.A. (2015). "Climate-induced larch growth response within the central Siberian permafrost zone". Environmental Research Letters, 10(12), 125009. doi.org/10.1088/1748-9326/10/12/125009.
Kukkonen, I.T., Suhonen, E., Ezhova, E., Lappalainen, H., Gennadinik, V., Ponomareva, O., … Drozdov, D. (2020). "Observations and modelling of ground temperature evolution in the discontinuous permafrost zone in Nadym, north‐west Siberia". Permafrost and Periglacial Processes, 31(2), 264-280. doi.org/10.1002/ppp.2040.
Matyshak, G.V., Goncharova, O.Y., Moskalenko, N.G., Walker, D.A., Epstein, H.E., & Shur, Y. (2017). "Contrasting Soil Thermal Regimes in the Forest-Tundra Transition Near Nadym, West Siberia, Russia". Permafrost and Periglacial Processes, 28(1), 108-118. doi.org/10.1002/ppp.1882.
Mekonnen, Z.A., Riley, W.J., Randerson, J.T., Grant, R.F., & Rogers, B.M. (2019). "Expansion of high-latitude deciduous forests driven by interactions between climate warming and fire". Nature Plants, 5(9), 952-958. doi.org/10.1038/s41477-019-0495-8.
Melnikov, E.S., Leibman, M.O., Moskalenko, N.G., & Vasiliev, A.A. (2004). "Active-Layer Monitoring in the Cryolithozone Of West Siberia". Polar Geography, 28(4), 267-285. doi.org/10.1080/789610206.
Myers-Smith, I.H., Kerby, J.T., Phoenix, G.K., Bjerke, J.W., Epstein, H.E., Assmann, J. J., … Wipf, S. (2020). "Complexity revealed in the greening of the Arctic". Nature Climate Change, 10(2), 106-117. doi.org/10.1038/s41558-019-0688-1.
Nitze, I., & Grosse, G. (2016). "Detection of landscape dynamics in the Arctic Lena Delta with temporally dense Landsat time-series stacks". Remote Sensing of Environment, 181, 27-41. doi.org/10.1016/j.rse.2016.03.038.
Osawa, A., Matsuura, Y., & Kajimoto, T. (2010). Characteristics of Permafrost Forests in Siberia and Potential Responses to Warming Climate (pp. 459-481). doi.org/10.1007/978-1-4020-9693-8_24.
Overland, J.E., Wang, M., Walsh, J.E., & Stroeve, J.C. (2014). "Future Arctic climate changes: Adaptation and mitigation time scales". Earth’s Future, 2(2), 68-74. doi.org/10.1002/2013EF000162.
Post, E., Alley, R.B., Christensen, T.R., Macias-Fauria, M., Forbes, B.C., Gooseff, M.N., … Wang, M. (2019). "The polar regions in a 2°C warmer world". Science Advances, 5(12), eaaw9883. doi.org/10.1126/sciadv.aaw9883.
Singh, V.P., Singh, P., Bishop, M.P., Björnsson, H., Haritashya, U.K., Haeberli, W., … Tranter, M. (2011). Encyclopedia of Snow, Ice and Glaciers. Springer Netherlands.
Tchebakova, N.M., Parfenova, E., & Soja, A.J. (2009). "The effects of climate, permafrost and fire on vegetation change in Siberia in a changing climate". Environmental Research Letters, 4(4), 045013. doi.org/10.1088/1748-9326/4/4/045013.
Thomas, H.J.D., Bjorkman, A.D., Myers-Smith, I.H., Elmendorf, S.C., Kattge, J., Diaz, S., … de Vries, F.T. (2020). "Global plant trait relationships extend to the climatic extremes of the tundra biome". Nature Communications, 11(1), 1351. doi.org/10.1038/s41467-020-15014-4.
Walker, M.D., Wahren, C.H., Hollister, R.D., Henry, G.H.R., Ahlquist, L.E., Alatalo, J.M., … Wookey, P.A. (2006). "From The Cover: Plant community responses to experimental warming across the tundra biome". Proceedings of the National Academy of Sciences, 103(5), 1342-1346. doi.org/10.1073/pnas.0503198103.
Wilmking, M. (2005). "Increased temperature sensitivity and divergent growth trends in circumpolar boreal forests". Geophysical Research Letters, 32(15), L15715. doi.org/10.1029/2005GL023331.
Yletyinen, J. (2019). "Arctic climate resilience". Nature Climate Change, 9(11), 805-806. doi.org/10.1038/s41558-019-0616-4.

Samuel Amos
Samuel Amos est diplômé du Master en sciences de l’environnement de l’Institut des sciences de l’environnement de l’Université de Genève. Il est titulaire du Certificat complémentaire de géomatique et d’un Bachelor en relations internationales du Global Studies Institute de l’Université de Genève, obtenu en 2017. Il s’intéresse aux politiques de l’environnement, aux changements climatiques ainsi qu’à la réduction des risques en cas de catastrophes. Dans le cadre de ses recherches il a étudié les avalanches de janvier 2018 comme événements extrêmes en combinant une approche dendrogéomorphologique et géomatique. En 2019 il a pris part à l’expédition interdisciplinaire Changing Arctic - Yamal au sein de l’équipe de recherche sur l’environnement.

Mathieu Logeais
Mathieu Logeais a obtenu un Master en management et bénéficie de quelques années d’expérience dans le domaine de la finance. Convaincu qu’il faut agir vite pour faire face au réchauffement de la planète, il a voulu se former et est retourné sur les bancs de l’université. Il a obtenu un Master en sciences de l’environnement ainsi qu’un Certificat complémentaire en géomatique de l’Institut des sciences de l’environnement de l’Université de Genève. C’est en participant au programme Changing Arctic et en récoltant les échantillons d’arbres qu’il a souhaité approfondir ses connaissances en dendroclimatologie, en étudiant les arbres des hautes latitudes dans le cadre de son travail de master.


